「ウリ類炭疽病菌のケルヒリピートタンパク質は微小管に結合し、付着器形成に関与する」
A. Sakaguchi, T. Miyaji, G. Tsuji and Y.Kubo (2010) A
Kelch repeat protein Cokel1p associates with microtubules and is
involved in appressorium development in Colletotrichum orbiculare. Molecular Plant-Microbe Interactions 23: 103-111.
主著者は大学院生の坂口 歩君です
論文のメッセージ
2008年にCoKEL2(旧名ClaKEL2)について報告しました。糸状菌におけるケルヒリピートモチーフをもつタンパク質機能の解明として世 界に先駆けて出した論文です。今回の論文は遺伝子挿入変異株のスクリーニングにより獲得した病原性変異株の解析から明らかにした遺伝子機能に関するもので
す。実は2008年に報告したCoKEL2は今回の変異株の変異遺伝子のモチーフの保存性に基づいて遺伝子をクローニングしたもので、変異株としてはこち
らを先に分離していました。先に報告したCokel2タンパク質は細胞の成長端に局在し、この局在には微小管を通じたタンパク質の移動が関与しています。
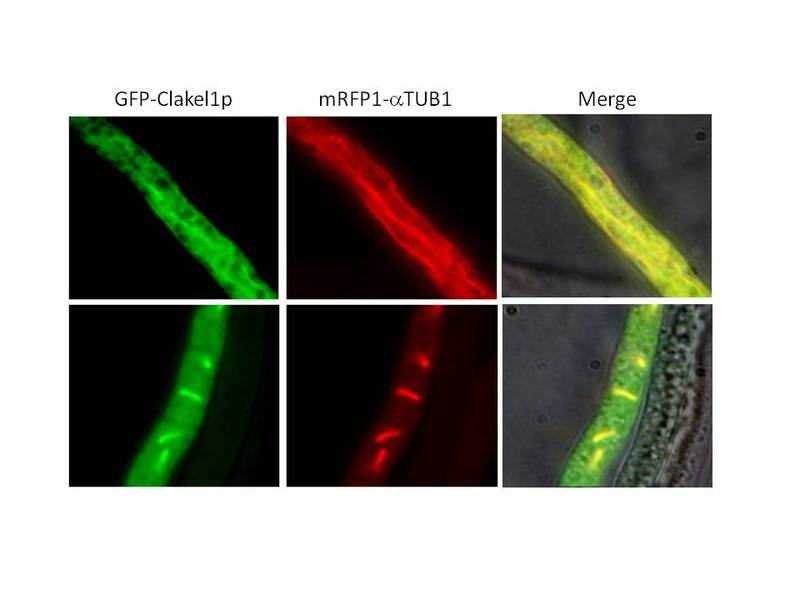
さて、この遺伝子のコードするタンパク質の局在はどうでしょうか?大学院生の坂口君が様々な観点から実験をデザインし、新しい知見を得ることに成功しまし
た。
1)Cokel1タンパク質の局在は観察した細胞によって異なります。核分裂過
程の細胞では核の微小管に結合しているところが観察されます。その他の細胞では細胞質の微小管に結合しているものや、局在が認められない細胞があります。
このことから局在が細胞のage, condition,あるいは細胞周期依存的に制御されていることが示唆されました。
2)病原菌の形態形成や病原性に対する影響は?cokel1変異株は形態的に異常な付着器を形成し、この付着器は宿主植物に対する感染能を欠いており、したがって、植物に対する病原性を欠いていました。
3)では付着器のどのような機能に欠損がみられるのでしょうか。一つは細胞壁の
健全性が影響を受け、細胞壁結合性染色剤に対する結合性や細胞壁分解酵素に対する感受性が変化していました。また、付着器の侵入能の発揮に必要な膨圧が野
生株に対して低下していました。このようなことから、付着器は十分な侵入能力を有しないと考えられました。
4)前回報告したCoKEL2は付着器の形態形成に関与しますが、想定されるシ
グナル伝達のバイパスの存在から、植物に対する病原性は維持されています。今回報告したCoKEL1は病原性に関与していることが示されました。
CoKEL1は病原性に関与することが示された初めてのケルヒモチーフをコードする遺伝子となります。
「ウリ類炭疽病菌のssd1変異株はMAPキナーゼを活性化することによりベンサミアーナタバコの基本的抵抗性を増強する」
S. Tanaka, N. Ishihama, H. Yoshioka, A. Huser, R. O'Connell, G. Tsuji,
S. Tsuge, and Y. Kubo (2009) The Colletotrichum orbiculare ssd1 mutant
enhances Nicotiana benthamiana basal resistance by activating a mitogen-activated
protein kinase pathway Plant Cell 21:2517-2526.
名古屋大学、マックスプランク研究所との共同研究です。(主著者は大学院生の田中茂幸君です)
論文のメッセージ
1)2007年にMolecular Microbiologyに発表した論文で
は、出芽酵母のSSD1という細胞壁の形成に関係している制御遺伝子のオルソログ遺伝子をノックアウトしたウリ類炭疽病菌とイネいもち病菌は宿主の病原菌
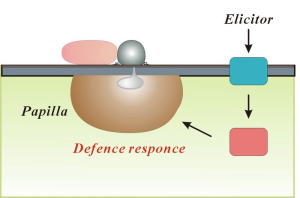
に対する防御反応(キュウリではパピラ形成、イネでは細胞死を伴う応答)を強く誘導すること。また、アブシジン酸処理やヒートショックで宿主の防御系を乱
すと変異株は感染性を復帰することを示しました。このことから、ssd1遺伝子変異は野生型株に比較して強い防御応答の誘導をもたらし、このことにより感
染性が低下することを示しました。
2)今回の論文では誘導される防御応答系にはどのような経路が関与するかについて、遺伝学的に明らかに
すること、そしてその防御応答系が実際にssd1変異によって強く誘導されているのかどうかを検証することをテーマとして行いました。そのため研究ではウ
リ類炭疽病菌の宿主として遺伝子サイレンシングの手法が安定して適用できるベンサミアーナタバコを用いました。
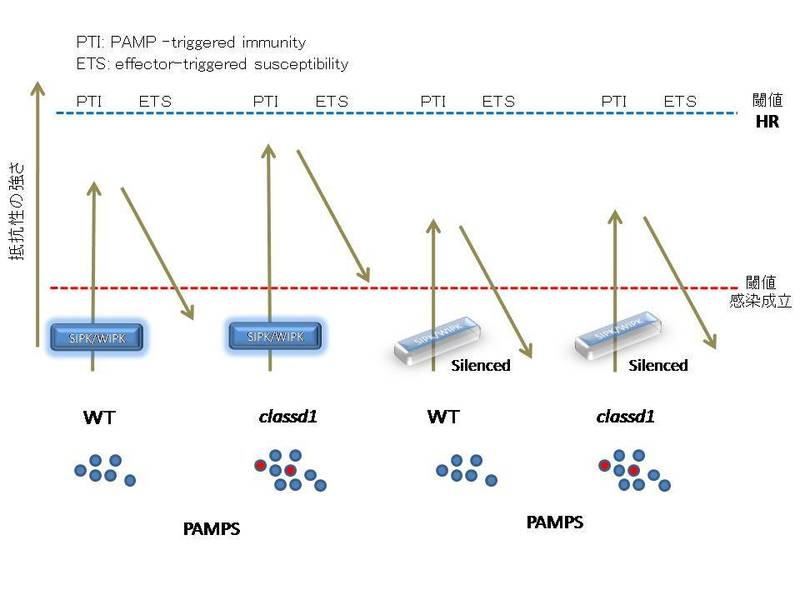
3)防御応答に関与すると
考えられる様々な遺伝子のサイレンシングしたところWIPK/SIPKキナーゼの二重サイレンシング、およびその上流のMEK2キナーゼのサイレンシング
が顕著な感染誘導を示しました。このことからこの経路が基本的抵抗性に関与していることを示しました。また、その他、R遺伝子特異的な抵抗性に関与する因
子などはサイレンシングによる効果は認めれませんでした。
4)では実際にssd1変異株が野生型よりも強いキナーゼ誘導活性をもっているかどうかを調べたところ、WIPK、SIPK活性がより強いことを免疫沈降キナーゼアッセイで明らかにしました。
5)ではssd1の変異による病原菌細胞表層の変化がキナーゼの活性を高めているかどうか。そのことを確認するために、熱処理して殺した胞子、さらにタンパク質分解酵素処理などいくつかの実験区を設定し、細胞表層の変化が活性に対応していることを示しました。
6)これらの結果から先にこのブログでも紹介したzigzagモデルとssd1変異との関係を論文で提唱しました。植物病原糸状菌の細胞表層構造に関わる変異で植物体との実際の相互作用の場面、すなわちin vivoで植物の防御応答のレベルに影響を与え、かつそれが感染の可否を決定する事例の初めての報告であると思われます。
7)これからの課題は?
・ssd1変異によって防御応答の誘導活性を示す分子種はどのような実体なのか?
・植物側の受容体は何なのか?
・そしてその受容体の下流で制御されるシグナル経路はどのようなものなのか?
・ssd1変異で誘導される基本的抵抗性を抑えるエフェクター因子を想定するならば、それはどのようものなのか。
これらを解明すべく、実験を進めています。さらに共同研究も進めていきたいと考えています。
A. Sakaguchi, T. Miyaji, G. Tsuji and Y. Kubo (2007) Kelch-repeat protein
Clakel2p and calcium signaling control appressorium development in Colletotrichum
lagenarium Eukaryotic Cell (2008) 7:102-111
1)
ウリ類炭疸病菌の病原性が低下する変異株を調べる中で、付着器が異常な形態をする変異株を分離しました。その変異株の変異遺伝子を調べてみますとケルヒリ
ピートと言われるモチーフをもつタンパク質をコードしていることがわかりました。このモチーフはタンパク質の相互作用に関与しています。ウリ類炭疸病菌で
は侵入器官の機能に関与していることから、このモチーフをもっている遺伝子の機能を調べてみようというのが研究の発端です。
2) 研究では2つのケルヒ‐リピートタンパク質遺伝子を単離することができました。今回の論文はClaKEL2遺伝子に関しての研究をまとめたものです。この遺伝子の変異株は次のような性質を示しました。
・付着器の形態が異常になる。
・セロファン膜を用いた侵入試験では異常付着器を形成し、侵入能力を失っている。
・しかし、植物上では正常な付着器を形成し、病原性を示す。
3) こうした変異株の性質から、ClaKEL2は形態分化に関連する遺伝子であること。しかしながら、植物の上ではClaKEL2を介さないバイパスが存在し、付着器形成を誘導することが考えられました。
4)
そうしますと、植物表面でないところでも植物の表面の成分を与えてやると正常な付着器形成が誘導されるはずです。実験ではキュウリ表面の滲出液を加えて変
異株の付着器形成を見てみると正常な付着器が回復しました。このことは、植物の成分を感知するシグナル伝達経路が存在していることを示しています。
5)
では、もしそうならcAMPやジアシルグリセロールやカルシウムといった細胞内2次メッセンジャーといわれる分子が付着器形成を誘導するかもしれません。
実験結果はカルシウムが付着器形成を回復する効果があること、さらにカルシウムチャンネルブロッカーといわれる阻害剤を用いるとその効果が失われることか
ら、カルシウムシグナル伝達の関与が強く示唆されました。
6)
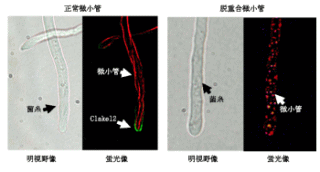
さらに、研究ではClaKEL2がコードするタンパク質の細胞での局在も調べました。ClaKEL2は分裂酵母のTea1という遺伝子に最も高い相同性を
示します。この遺伝子は細胞極性形成に関与していることが分裂酵母で示されています。GFP融合タンパク質で可視化したところ、細胞の成長端に局在してい
ることがわかりました。また、この局在は菌糸生育では微小管の働きに依存していることもわかりました。
7)以上から付着器形成には細胞の成長極性に関わるClaKEL2が関与していること。一方、ウリ類炭疸病菌が植物に感染するときにはClaKEL2を介しない植物に特徴的なシグナルを利用していることがわかりました。。 |
|